Tubuh manusia, tanpa berlebihan, dapat digolongkan di antara sistem biologis paling kompleks di planet ini. Tubuh kita terdiri dari milyaran sel, banyak organ dan sistem. Tetapi semua kemegahan dan keragaman ini muncul karena peleburan hanya dua sel - sperma dan telur. Saya pikir tidak perlu menjelaskan bagaimana pemupukan terjadi dan apa yang diperlukan untuk ini (petunjuk - bangau dan kubis tidak ada hubungannya dengan itu). Namun berikut beberapa aspek kehidupan sperma yang selama bertahun-tahun masih belum jelas. Ilmuwan dari Universitas Bristol, dengan menggunakan teknik modern mikroskop tiga dimensi, mampu melihat pergerakan sperma dengan cara yang sebelumnya tidak mungkin dilakukan. Bagaimana dan dengan cara apa sperma bergerak, kita belajar dari laporan para ilmuwan. Pergilah.
Dasar penelitian
Terlepas dari kenyataan bahwa sperma terlibat dalam penciptaan kehidupan jauh sebelum munculnya metode ilmiah, jalur mereka dalam literatur ilmiah dimulai baru-baru ini - pada tahun 1677. Mahasiswa kedokteran Johann Gam berbagi pengamatannya dengan kolega dan temannya Anthony van Leeuwenhoek (1632 - 1723), yang, pada gilirannya, memeriksa dan menjelaskan secara rinci "hewan mani" (sebagaimana ia disebut spermatozoa).
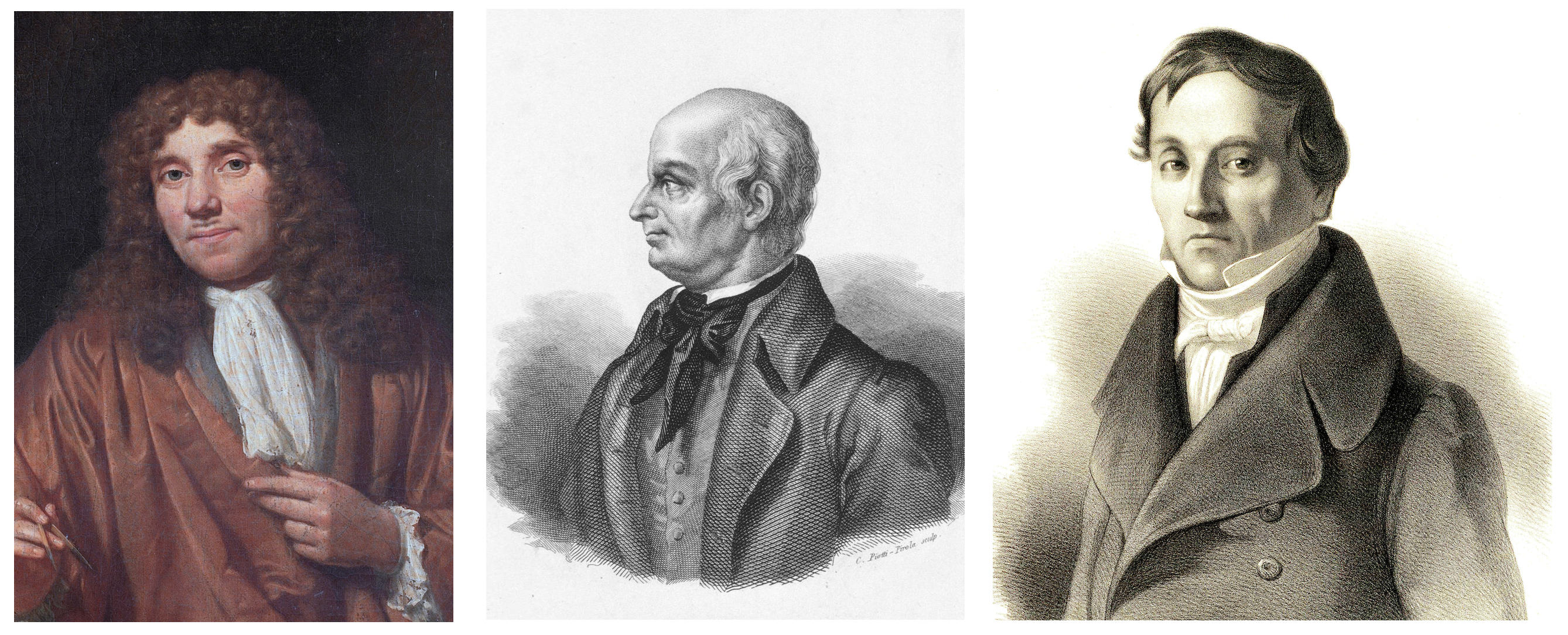
Anthony van Leeuwenhoek / Lazzaro Spallanzani / Carl Ernst von Baer
Leeuwenhoek mengemukakan bahwa sel-sel yang tidak biasa ini terlibat dalam pembuahan, tetapi teorinya, meskipun benar, ditolak oleh komunitas ilmiah. Untuk waktu yang lama diyakini bahwa sperma adalah parasit, dan hanya cairan mani yang terlibat dalam pembuahan.
Hanya hampir seratus tahun kemudian, pada tahun 1786, fakta partisipasi sperma dalam pembuahan dibuktikan oleh Lazzaro Spallanzani (1729 - 1799) dalam karyanya Experiencias Para Servir a La Historia de La Generación De Animales y Plantas. Namun, penjelasannya untuk proses itu sendiri agak kabur: dia percaya bahwa sel telur sudah merupakan awal dari organisme baru, dan sperma hanya dibutuhkan untuk mengaktifkan proses pertumbuhan.
Istilah yang sama "sperma" diperkenalkan pada awal abad ke-19 oleh Karl Ernst von Baer (1792 - 1876).
Apa pun yang dipikirkan para ilmuwan beberapa abad yang lalu, sperma memiliki fungsi yang sangat jelas, yang implementasinya disediakan oleh sejumlah alat khusus. Tugas utama sperma manusia adalah melewati saluran kelamin perempuan, menemukan sel telur dan mentransfer materi genetik laki-laki kepadanya.

Struktur sperma
Sel reproduksi laki - laki tidak dapat membanggakan dimensi, karena merupakan yang terkecil dalam tubuh manusia (tidak termasuk ekor): dimensi kepala 5,0x3,5x2,5 mikron (panjang x lebar x tinggi), panjang bagian tengah 4,5 mikron, dan panjang ekor - 45 mikron.
Pada saat yang sama, ukurannya yang kecil bukanlah suatu kerugian, tetapi aspek yang bijaksana untuk meningkatkan kecepatannya. Dalam proses pematangan sperma, nukleusnya (membawa satu set kromosom) menjadi lebih padat, sebagian besar sitoplasma dibuang, dan hanya organel terpenting yang tersisa di dalam sel.
Flagel dapat disebut elemen terpenting kedua dari sperma setelah nukleus, mis. ekornya. Karena berkat dialah pergerakan sel ini di sepanjang saluran genital seorang wanita dilakukan. Lucu juga bahwa lingkungan vagina sangat merusak sel germinal pria, tetapi air mani mengurangi sebagian efek negatif pada sel sperma. Tingkat pH di dalam saluran genital wanita memungkinkan sperma bergerak menuju rahim, di mana lingkungan yang lebih menguntungkan menanti mereka.
Sebelumnya, sperma diyakini bergerak maju karena gerakan simetris flagelnya dari sisi ke sisi.
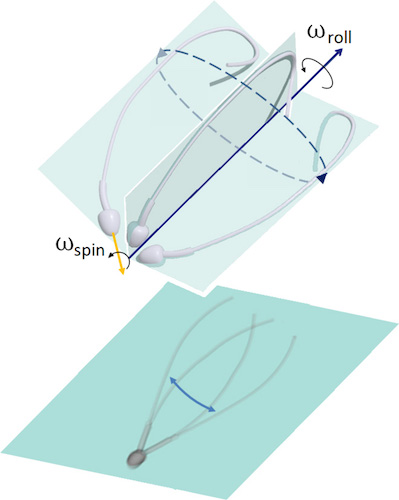
Gambar # 1: asimetri gerakan flagela dalam 3D (atas); proyeksi planar gerakan flagel, menciptakan ilusi optik simetri bilateral dalam mikroskop 2D (bawah).
Pernyataan ini diungkapkan bahkan di zaman Levenguk. Ini juga mengarah pada idealisasi simetris bentuk gelombang dalam tiga dimensi, sering dianggap sebagai spiral kerucut, mirip dengan pembuka botol yang mengembang.
Karena mikroskop dua dimensi, banyak pengamatan telah ditafsirkan secara tidak akurat, dan terkadang salah sama sekali. Pernyataan mengenai kesimetrian sayap flagel bertentangan dengan banyak pengamatan yang menunjukkan asimetri struktural dalam kerangka flagel itu sendiri.
Jika kepakan flagel sperma selama gerakan masih asimetris, lalu bagaimana simetri gerakan flagel dari sisi ke sisi dan pergerakan sel ke arah depan tercapai? Pertanyaan inilah yang menjadi pertanyaan utama dalam studi ini.
Untuk mendapatkan jawabannya, para ilmuwan membandingkan pengamatan molekuler dan mikroskopis yang menunjukkan bahwa sperma manusia menggunakan kontrol asimetris dan anisotropik untuk mengatur kepakan flagela. Dengan kata lain, simetri diwujudkan karena asimetri: efek dari "presesi puncak" terjadi ketika kepala berputar ("cairan pengeboran", kata-kata penulis) secara bersamaan dan terlepas dari bagaimana flagel sperma berputar di sekitar sumbu gerak.
Hasil penelitian
Gerakan cepat flagela sperma manusia telah direkam dengan resolusi spasial-temporal yang tinggi dalam 3D. Dua kelompok spermatozoa yang mengambang bebas dalam cairan dengan viskositas rendah diperiksa: spermatozoa yang mengapung di samping penutup kaca (ditempatkan di atas spesimen selama mikroskop) dan menjauh darinya.
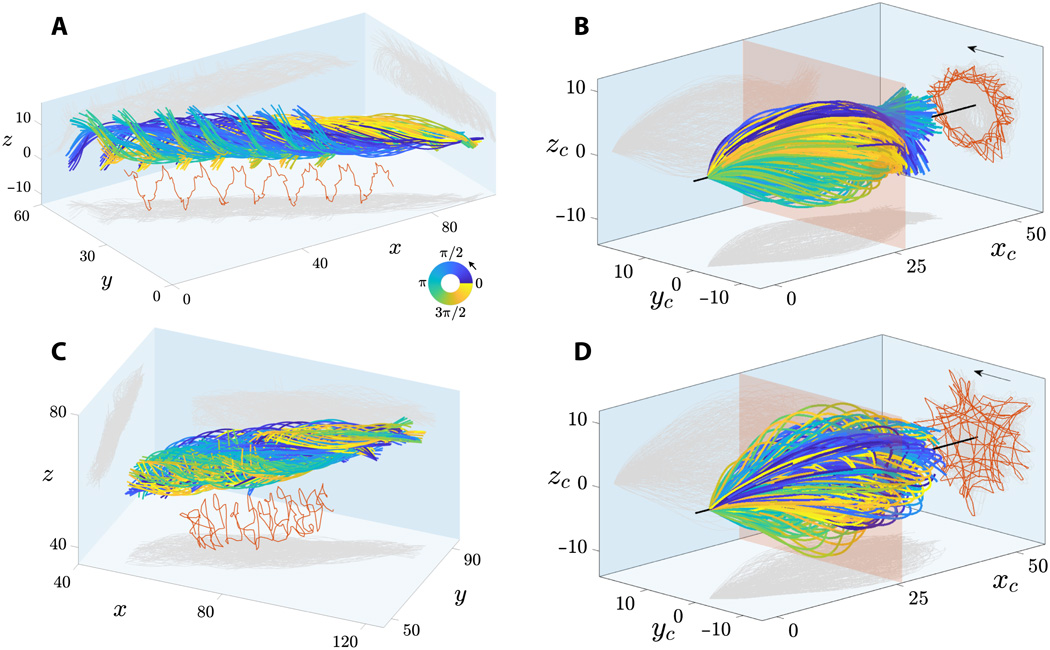
Gambar №2
Pada 2A dan 2C menunjukkan sapuan flagela sperma yang mengambang di dekat dan jauh dari kaca penutup. Flap flagel dicirikan oleh karakteristik gerakan berguling di sekitar arah pergerakan sperma.
Rotasi gabungan dan gerakan translasi dari flagel sperma mengarah ke lintasan spiral dari titik tengah flagel dengan chirality yang diawetkan (ditandai dengan warna merah pada 2Adan 2C ). Spermatozoa menunjukkan rotasi dua arah di sekitar porosnya: semua sel yang mengambang bebas (28 buah) berputar berlawanan arah jarum jam jika dilihat dari ujung depan (panah pada 2B dan 2D ) dan hanya 2 sel yang diputar searah jarum jam (mereka tidak bergerak maju karena hambatan dalam perjalanan).
Mikroskopi spermatozoa mengambang di samping penutup kaca (sesuai dengan 2A ).
Semua spermatozoa (30 sel) bergerak serupa ke bagian atas yang berputar sebelumnya, di mana rotasi kepala di sekitar sumbu longitudinal sel sperma (ω spin ) terjadi secara bersamaan dan terlepas dari bagaimana flagel berputar relatif terhadap sumbu gerak (ω roll ).
Pada 2B dan 2D menunjukkan flagel yang mengepak dari sistem koordinat * bersamaan , yaitu titik pandang, bergerak bersama dengan sperma, tetapi bukan gerakan berputar di sekitar porosnya. Ini menunjukkan bahwa goresan flagela sangat simetris baik di bidang ( xy ) maupun di luar bidang (z ) arah, yang sesuai dengan pengamatan dalam 3D.
Kerangka acuan yang menyertai * adalah kerangka acuan yang terkait dengan tubuh yang bersangkutan pada saat tertentu. Tubuh di dalam sistem ini tidak bergerak. Sebagai contoh, elevator yang jatuh bebas adalah kerangka acuan yang menyertai benda yang jatuh bebas di dalamnya, tetapi sistem bumi bukanlah sistem yang terkait dengan benda di elevator.Proyeksi lintasan tengah flagel (garis merah dalam 2B dan 2D ) menunjukkan susunan pola geometris yang luar biasa, dari bintang berputar hingga segitiga, persegi, dan pola lingkaran dengan simetri kutub. Pola flagela tidak beraturan yang ditunjukkan dalam 2D untuk spermatozoa yang mengapung dari penutup penutup juga diamati untuk sperma yang mengambang di sebelah penutup penutup. Variabilitas dalam pola ini dapat disebabkan oleh ketidaksesuaian dalam kelambatan fase antara komponen dalam bidang dan di luar bidang sayap untuk setiap sel, yang meningkat dalam garis waktu gerakan bergulir. Akibatnya, pola flagela bukanlah ciri pembeda sel yang mengapung di dekat dan jauh dari penutup kaca.
Mikroskopi dan pemodelan flagel yang menunjukkan rotasi kepala dan flagel di sekitar sumbu gerak.
Amplitudo gelombang 3D dicirikan oleh kurva amplop berbentuk peluru yang simetris, berlawanan dengan spiral kerucut (mirip dengan pembuka botol yang mengembang), yang sering dijelaskan dalam literatur. Spermatozoa yang melayang jauh dari penutup kaca memiliki bentuk gelombang yang lebih simetris daripada sel yang mengapung di samping kaca. Jadi, penutup kaca yang berdekatan merupakan sumber asimetri yang lemah untuk pengepakkan flagela.
Spermatozoa yang mengapung di sebelah kaca penutup memiliki sudut serang * -7 ° yang tetap, dengan orientasi rata-rata flagel sperma mengarah ke penutup kaca penutup.
* — ( ) , .

Gambar # 3
Gambar di atas menunjukkan perbandingan goresan flagela di bingkai pendamping (baris atas) dan bingkai bergulir yang menyertainya (baris bawah).
Gambar 3E menunjukkan sifat sebenarnya dari hentakan flagela dilihat dari perspektif sperma, tanpa gerakan mengambang atau berguling.
Kerangka referensi bergulir ( 3E ) yang menyertai menunjukkan bahwa flap flagela bersifat anisotropik, mis. karakteristik gelombang di setiap arah transversal (tegak lurus dengan bidang sayap), berlabel " bidang b " (bidang biru) dan " bidang z " (bidang merah), sangat berbeda.
Jika kita membandingkan area abu-abu terang pada bidang biru dan merah di 3E, maka dapat dilihat bahwa kepakan tersebut sangat asimetris pada bidang b dan dicirikan oleh kerusakan simetri kiri-kanan, yang mengingatkan pada bentuk-C.
Pengamatan ini sangat kontras dengan pola simetris yang diamati dalam kerangka acuan bergerak di 3A .
Bentuk gelombang flagela relatif terhadap kerangka acuan tetap laboratorium (x, y, z).
Analisis komponen utama (PCA) dari bentuk gelombang tiga dimensi telah memungkinkan para ilmuwan untuk menguraikan gelombang flagela menjadi beberapa "mode bentuk dasar," yang di sini disebut sebagai mode PCA. Bentuk gelombang 3D dapat direkonstruksi dengan akurasi yang baik hanya dengan dua mode bentuk gelombang, seperti yang terlihat pada 3B dan 3F .
Pada 3C, terlihat bahwa dua mode PCA pertama memiliki bentuk yang identik, hingga rotasi 90 °, sehingga menangkap bentuk spiral yang ramping yang disebabkan oleh penggulungan sperma.
Pada 3G , bagaimanapun, bentuk C asimetris internal sepenuhnya terdeteksi hanya oleh mode PCA pertama. Mode PCA kedua di 3G(Disorot dalam warna oranye) memperkenalkan deviasi kecil tegak lurus ke mode PCA pertama (disorot dengan warna biru), menunjukkan bahwa bentuk gelombang dapat diuraikan menjadi dua arah gelombang melintang independen yang anisotropik di alam.
Analisis Fourier dari sapuan memungkinkan untuk merekonstruksi pergerakan flagel menggunakan hanya dua mode Fourier pada 3D dan 3H . Singkatnya, setiap sinyal ayunan umum dapat didekati dengan jumlah sederhana dari dua fungsi:
f r (s, t) ≈ f 0 (s) + | f 1 (s) | sin (ωt + φ (s))Fungsi pertama f 0 (s) tidak bergantung pada waktu, ini disebut "mode statis", dan ini memperbaiki asimetri sinyal rata-rata sepanjang busur flagel.
Fungsi kedua adalah gelombang perjalanan sinusoidal yang disebut "mode dinamis", berosilasi pada frekuensi yang ditangkap oleh puncak pertama dari spektrum daya sinyal.
Amplitudo dan modulasi fase dari gelombang yang bergerak sepanjang flagel adalah sama, | f 1 (s) | dan φ (s) = arg (f 1 (s)).
Dengan demikian, fase membawa informasi tentang karakteristik gelombang berjalan. Misalnya, jika fasa φ (s) tidak berubah sepanjang s, maka sinyalnya bukanlah gelombang berjalan, melainkan gelombang berdiri.
Oleh karena itu, mode statis menangkap ketidakcocokan bentuk gelombang apa pun. Dalam hal ini, garis lurus hitam dalam 3D mencerminkan kesimetrian sapuan di bidang xy dan xz dalam bingkai comoving. Sebaliknya, dalam kerangka referensi bergulir pendamping ( 3H ), mode statis (kurva hitam) dicirikan oleh amplitudo asimetris yang besar, yang mengingatkan pada C terbalik.
Bentuk gelombang flagel relatif terhadap kerangka acuan yang menyertainya.
Bentuk gelombang flagel relatif terhadap kerangka referensi bergulir yang menyertainya.
Modus dinamis dalam bingkai pendamping (kurva merah) dalam 3D memiliki amplitudo yang besar dan sangat simetris di bidang xy dan xz karena perputaran sperma.
Tetapi mode dinamis dalam kerangka referensi bergulir (kurva merah) pada 3H memiliki amplitudo bentuk gelombang yang berkurang dan arah perjalanan yang disukai.
Rekonstruksi Fourier dari bentuk gelombang dilakukan dengan menjumlahkan mode statis dan dinamis (grafik di tengah pada 3D dan 3H ), yang sesuai dengan pengamatan asli (grafik di sebelah kanan dalam 3Ddan 3H ).
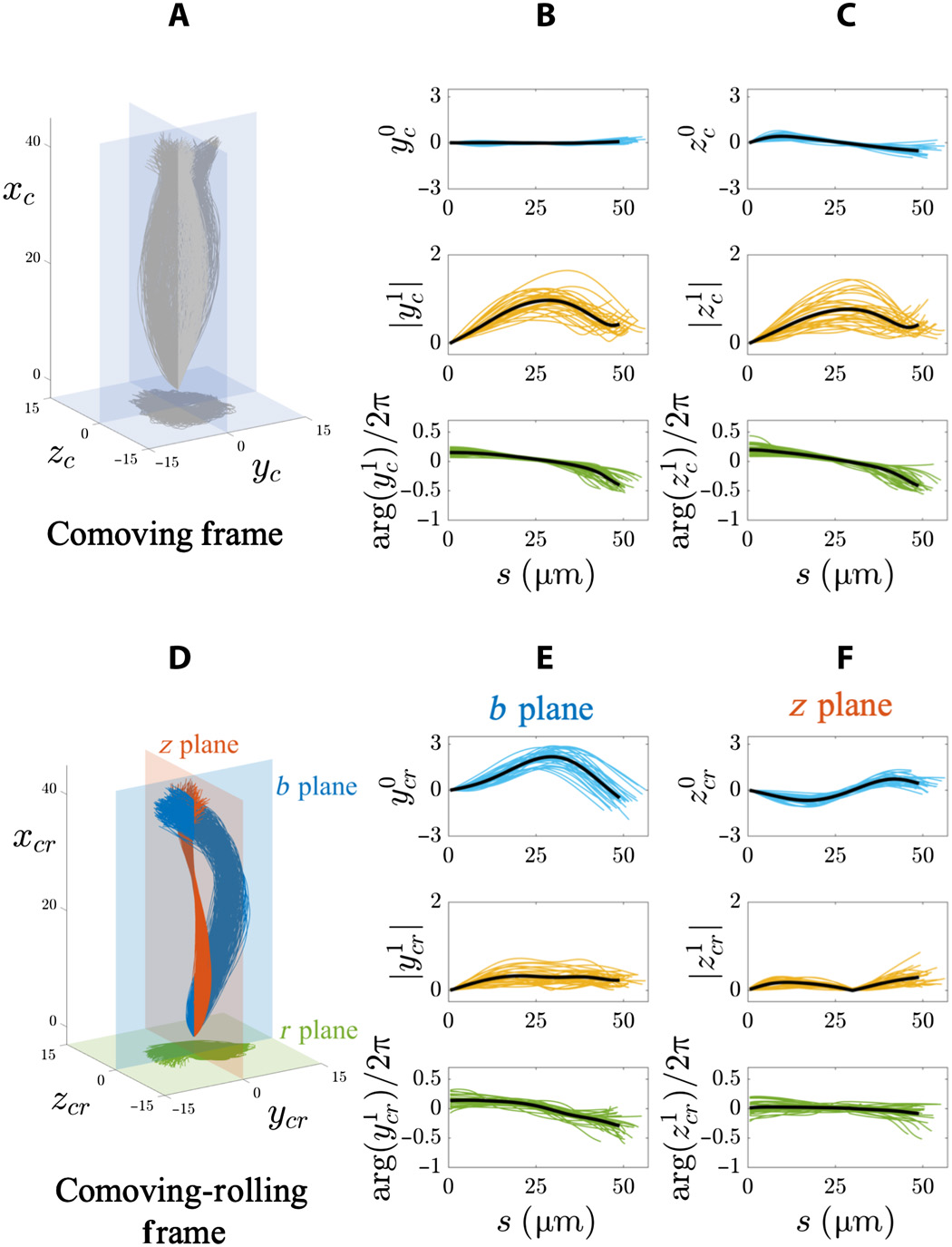
Gambar # 4
Grafik di atas menunjukkan hasil analisis Fourier dari flap flagela 3D pada populasi sperma yang mengambang bebas (20 sel di sebelah penutup penutup dan 8 sel jauh dari kaca). Dalam kerangka acuan yang menyertai ( 4A - 4C ), amplitudo mode statis di kedua arah (y c dan z c ) sangat kecil (baris atas grafik pada 4B dan 4C ) karena simetri sapuan.
Selain itu, amplitudo (baris tengah) dan fase (baris bawah) dari mode dinamis (y c dan z c ) pada 4B dan 4C.memperbaiki simetri transversal dan melebihi isotropi dalam kerangka acuan ini karena fakta bahwa spermatozoa berputar di sekitar sumbu gerak.
Karakteristik gelombang berjalan untuk kedua koordinat (y c dan z c ) adalah sama untuk semua sel yang mengambang bebas: frekuensi 4 Hz, panjang gelombang 100 μm, dan kecepatan gelombang 400 μm / s.
Dalam sistem referensi bergulir ( 4D - 4F ), rezim statis yang berbeda di bidang b dan z menunjukkan bahwa ayunan masih memiliki anisotropi dalam populasi sel mengambang bebas ( 4E dan 4F ). Pesawat
mode statis b (y cr) sangat asimetris dan bias ke arah nilai positif (baris atas grafik oleh 4E ). Tetapi untuk bidang z (z cr ), ia berosilasi secara simetris sepanjang busur secara sinusoidal (baris atas di 4F ).
Amplitudo rezim dinamis pada bidang b (y 1 cr ) meningkat hingga mencapai dataran tinggi (baris tengah pada 4E ), sedangkan rezim dinamis pada bidang z (z 1 cr ) nonmonotonik sepanjang busur (baris tengah pada 4F ). Karakteristik gelombang perjalanan y cr ( bbidang) adalah: frekuensi 8 Hz, panjang gelombang 145 μm dan kecepatan gelombang 1120 μm / s. Ciri-ciri gelombang perjalanan z cr ( bidang z ): frekuensi 6 Hz, panjang gelombang 1526 μm, dan kecepatan 5174 μm / s.
Perubahan kecil dalam fase pada jarak yang jauh di sepanjang flagel membutuhkan kecepatan perambatan gelombang yang sangat tinggi. Akibatnya, osilasi bidang z benar-benar berperilaku seperti gelombang berdiri yang berdenyut dalam waktu.
Mikroskopi spermatozoa non-propelling dan rolling.
Totalitas data di atas menunjukkan bahwa pengepakkan flagela menggunakan dua pengontrol transversal anisotropik koaktif, yang tidak terlalu berbeda dengan gelombang elektromagnetik yang berjalan. Namun, setiap gelombang geser (y cr , z cr ) adalah jumlah dari mode statis dan dinamis: gelombang perjalanan asimetris sepanjang bidang b (biru pada 4D ) dan gelombang berdiri simetris pada bidang z (merah pada 4D ).
Anehnya (tapi tidak mengherankan) bahwa keberadaan kaca penutup di dekat lintasan sel sperma juga berpengaruh pada amplitudo perambatan gelombang.

Gambar No. 5
Kaca mengurangi amplitudo perambatan gelombang karena interaksi hidrodinamik antara flagel dan permukaan keras kaca penutup.
Dalam bingkai pendamping ( 5A dan 5B ), amplitudo dari kedua mode dinamis (y c , z c ) menurun menjelang akhir flagel sperma dekat penutup penutup (grafik di tengah), sedangkan mode statis tetap tidak berubah (grafik puncak).
Modus dinamis z c hanya sedikit kurang dari y c (kurva biru) di plot tengah 5A dan 5B . Ini kontras dengan profil yang simetris dan tidak berubah dari keduanya (yc , z c ) mode dinamis untuk spermatozoa yang terdeteksi jauh dari kaca (kurva merah pada grafik di tengah 5A dan 5B ).
Jika kita memperhatikan sistem referensi gelinding yang menyertainya, maka sifat efek kaca cukup anisotropik, karena hanya mempengaruhi satu bidang gelinding, yaitu. di pesawat b ( 5C ).
Slip penutup mempengaruhi mode y cr statis dan dinamis (grafik di atas dan di tengah pada 5C ). Tetapi bidang z (z cr ) tetap tidak berubah (grafik di atas dan tengah di 5D ).
Bentuk mode statis dalam sistem referensi bergulir pendamping adalah sama di antara semua spermatozoa dan mendefinisikan spiral tangan kanan di luar pusat yang ditunjukkan oleh h (s) dan ditunjukkan oleh kurva hitam pada 5E . Proyeksi spiral ke bidang bergulir (bidang hijau) adalah spiral berlawanan arah jarum jam yang tidak memiliki simetri kutub, yaitu. offset ke satu sisi (proyeksi abu-abu pada 5E ).
Spiral mode statis sangat mirip dengan spiral logaritmik yang sering ditemukan di alam. Namun, dalam kasus ini, jari-jari spiral berubah secara non-monoton, meningkat / menurun pada tingkat yang lebih cepat daripada spiral logaritmik di tempat lain di alam.
Spiral logaritmik kanan h (s) pada 5Edapat diekspresikan dalam radius dan nada ( 5G ), yang meluruh secara eksponensial sepanjang flagel. Setiap perubahan tanda pada h y atau h z menyebabkan spiral beralih ke rotasi searah jarum jam, sehingga menciptakan spiral kidal. Semua spermatozoa mengambang bebas menghasilkan spiral berlawanan arah jarum jam. Hanya dua spermatozoa yang memiliki spiral searah jarum jam, meskipun dengan bentuk yang identik dengan 5E . Kedua sel inilah yang tidak dapat berenang ke depan karena beberapa rintangan di jalurnya, tetapi mereka dapat terus berputar di sekitar poros puntirnya sendiri.
Dalam semua kasus, rotasi heliks berkorelasi dengan arah penggulungan sperma sebagai berikut: heliks berlawanan arah jarum jam untuk rotasi searah jarum jam (dilihat dari ujung belakang) dan heliks searah jarum jam untuk rotasi berlawanan arah jarum jam.
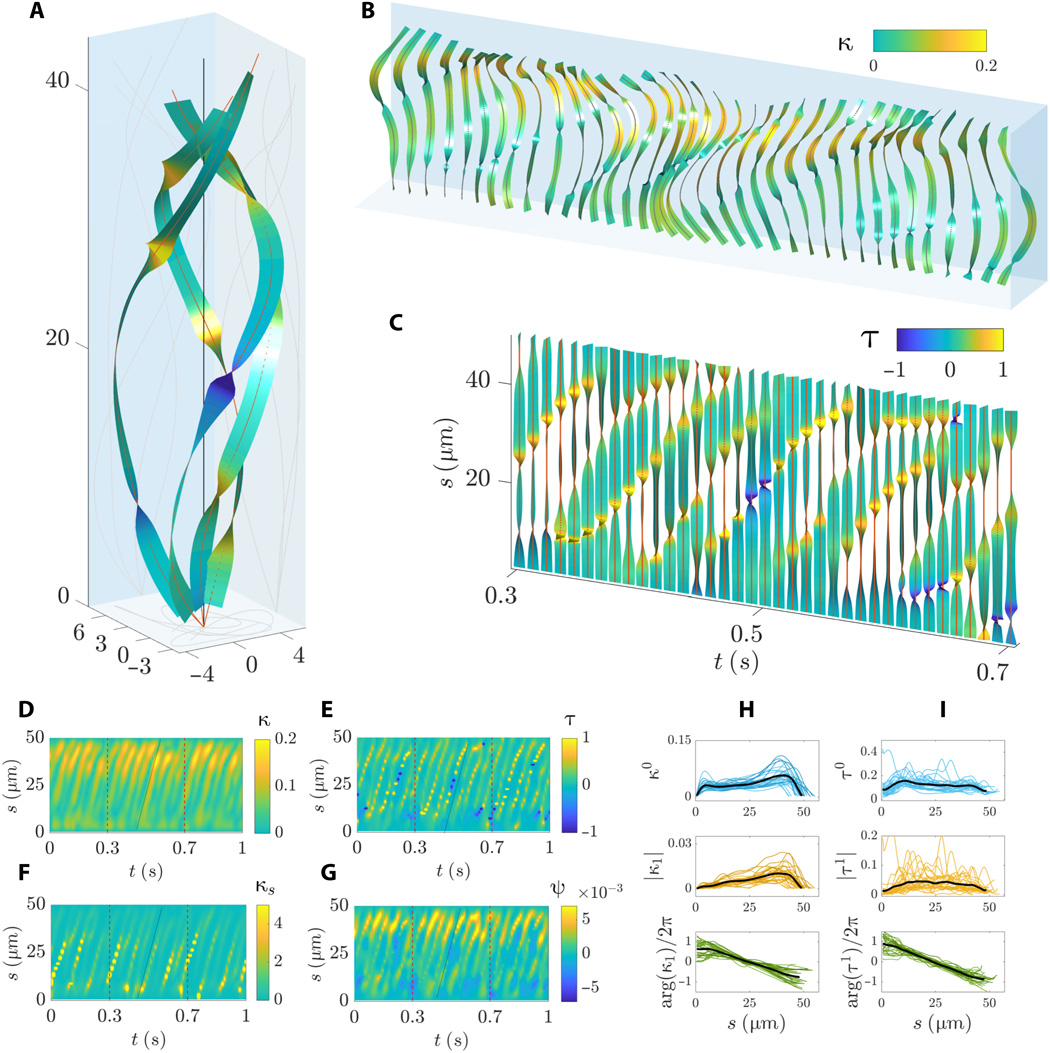
Gambar # 6
Grafik 6A dan 6B menunjukkan urutan kompleks gelombang berjalan saat flagel berputar di sekitar porosnya. Gelombang lentur merambat secara linier di sepanjang flagel dengan amplitudo non-monotonik sepanjang busur, ditandai dengan peningkatan tajam di daerah tengah dan distal.
Puntiran gelombang ditandai dengan tikungan tajam di sepanjang busur ( 6B dan 6C) dengan belokan positif dan negatif secara bersamaan. Bentuk spiral dari garis tengah flagel mengalami fenomena di mana daerah chirality berlawanan di sepanjang flagel hidup berdampingan. Namun, bagian-bagian flagel dengan kiral yang berlawanan bergerak selama pengepakkan ( 6C dan 6E ). Gelombang torsi yang bergerak merambat dengan kecepatan yang sama dengan gelombang kelengkungan.
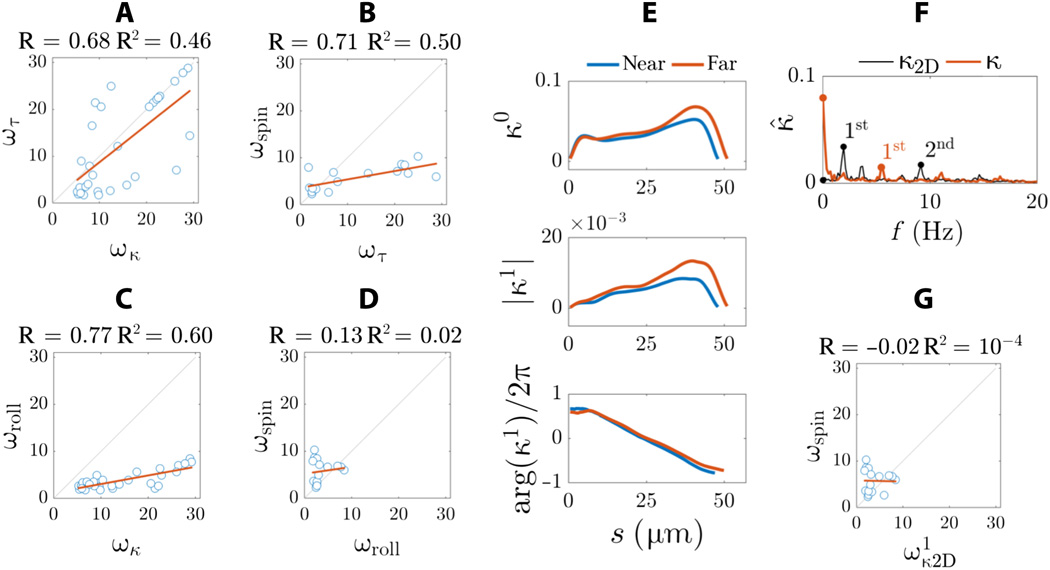
Gambar # 7
Untuk lebih memahami perbedaan antara hasil mikroskop 3D yang diperoleh dalam penelitian ini dan hasil mikroskop 2D yang sering dijelaskan dalam literatur, ilmuwan membuat proyeksi 2D dari bentuk gelombang 3D.
Regim statis kelengkungan dua dimensi sangat tidak signifikan ( 7F). Dalam 2D, asimetri intrinsik dari bentuk gelombang tidak dapat dideteksi. Sebaliknya, spektrum frekuensi dicirikan oleh dua puncak frekuensi (penanda hitam di 7F ) daripada satu puncak frekuensi utama yang terlihat untuk kelengkungan tiga dimensi (kurva merah di 7F ).
Untuk pengenalan yang lebih mendetail tentang nuansa penelitian ini, saya sarankan Anda melihat ke dalam laporan para ilmuwan dan bahan tambahan padanya.
Epilog
Dalam karya ini, para ilmuwan dapat menunjukkan dalam praktiknya bahwa karena mikroskop tiga dimensi, dimungkinkan untuk mengetahui apa yang tidak akan diberikan oleh mikroskop dua dimensi - misalnya, untuk melihat perubahan arah spiral (perubahan chirality) selama osilasi flagela.
Pada pandangan pertama, sperma tampak seperti melayang ke depan, mengibas-ngibaskan ekornya. Namun, pemeriksaan rinci terhadap kinematika sel-sel ini menunjukkan bahwa proses ini jauh lebih rumit. Mengepakkan dan mengayunkan flagela, rotasi sel itu sendiri - semua ini memungkinkan sperma untuk bergerak maju.
Beberapa informasi ini sebelumnya tersedia, tetapi tidak semua detail telah dirinci karena keterbatasan mikroskop 2D. Memahami bagaimana pergerakan sperma dapat banyak membantu di bidang pengobatan reproduksi, kata para ilmuwan. Secara khusus, data baru akan memperluas kisaran parameter yang digunakan untuk menentukan sel kelamin yang tidak sehat dari pria, yang merupakan penyebab infertilitas pada sekitar setengah dari pasangan.
Bagaimanapun, semakin baik kita memahami fenomena dan proses tertentu, terutama yang terjadi di tubuh kita sendiri, semakin besar kemungkinannya untuk meningkatkan kualitas hidup kita.
Terima kasih atas perhatiannya, tetap penasaran dan semoga minggu kerja Anda bagus, guys. :)
Sedikit iklan
Terima kasih untuk tetap bersama kami. Apakah Anda menyukai artikel kami? Ingin melihat konten yang lebih menarik? Dukung kami dengan memesan atau merekomendasikan kepada teman, cloud VPS untuk pengembang mulai $ 4,99 , analog unik dari server level awal yang kami ciptakan untuk Anda: The Whole Truth About VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps dari $ 19 atau bagaimana membagi server dengan benar? (opsi tersedia dengan RAID1 dan RAID10, hingga 24 core dan hingga 40GB DDR4).
Apakah Dell R730xd 2x lebih murah di pusat data Equinix Tier IV di Amsterdam? Hanya kami yang memiliki 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 TV dari $ 199 di Belanda!Dell R420 - 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB - Dari $ 99! Baca tentang Bagaimana membangun infrastruktur bldg. sekelas dengan server Dell R730xd E5-2650 v4 dengan biaya € 9000 untuk satu sen?